Mientras que en las plantas se conocen varias series de formas que conducen de la organización unicelular a la pluricelular, en el Reino Animal se sabe muy poco sobre la transición entre protozoos y metazoos. Dicha transición no está documentada por fósiles y las formas recientes supuestamente intermedias tampoco nos ayudan demasiado.
En este campo de la transición pueden mencionarse, por una parte, a Proterospongia, coanoflagelado marino y planctónico que forma una masa gelatinosa con coanocitos en la parte exterior y células ameboides en el interior, y por otra al pequeño organismo marino Trichoplax adhaerens (Filo placozoos) que forma una placa cerrada por epitelio pavimentosos en la parte dorsal y cilíndrico en la parte central, y presenta en la cavidad interior células en forma de estrella; se reproduce por yemas flageladas y huevos. Otra forma sencilla de metazoo es Xenoturbella, que vive sobre los fondos fangosos del mar. Tienen algunos centímetros de largo y forma de hoja, una boca ventral que conduce a un estómago en forma de saco. Entre la epidermis y el intestino existe una capa de tejido conjuntivo con un tubo muscular longitudinal y células musculares en el mesénquima; en la parte basal de la epidermis existe un plexo nervioso y en la parte anterior presenta un estatocisto; produce óvulos y espermatozoides, éstos idénticos a los de diferentes metazoos primitivos. Su posición sistemática es incierta, habiéndose propuesto como miembro de un filo independiente (xenoturbélidos), a emplazar tal vez en la base de los deuteróstomos. Por lo que respecta a los mesozoos, ya no son considerados un estado de transición entre Protistas y metazoos; su modo de vida parásito parece que les condujo a una reducción y simplificación extremas a partir de vermes acelomados.
Por tanto, se debe recurrir a la morfología, fisiología y ontogenia comparadas de los metazoos para poder reconstruir esta etapa de la evolución. Los datos obtenidos con microscopía electrónica y análisis moleculares han apagado antiguas controversias sobre el origen de los metazoos. En este sentido, parece definitivamente rechazada la hipótesis sobre un origen polifilético; incluso los placozoos y los mesozoos, considerados a veces como originados directa e independientemente de los protistas, parecen a la luz de los nuevos datos claramente metazoos. Tres son las principales teorías sobre el origen de los metazoos:5
Teoría colonial. La teoría más aceptada es la que postula que los metazoos tuvieron un origen colonial a partir de los coanoflagelados, un pequeño grupo de Mastigóforos monoflagelados; algunos son individuales y otros coloniales. Dicha teoría se ve avalada tanto por datos moleculares (ARN ribosómico) como morfológicos (las mitocondrias y las raíces flagelares son muy semejantes en los metazoos y en los coanoflagelados, un cierto número de metazoos presenta células tipo coanocito, y los espermatozoides son uniflagelados en la mayor parte de ellos). Los seguidores de esta teoría incluyen el filo Choanozoa en el reino animal, en contraposición al resto de animales, los metazoos. El antecesor de los metazoos, sería una colonia hueca y esférica de dichos flagelados; las células sería uniflageladas en su superficie externa; la colonia poseería un eje anteroposterior, nadando con el polo anterior hacia delante; entre las células somáticas existirían algunas células reproductoras. Este estado hipotético se ha denominado blastaea, y se cree que es el reflejo del estado de blástula que se produce en el desarrollo de todos los animales. Por tanto, esta teoría considera que los animales han evolucionado de protozoos flagelados. Sus parientes vivos más cercanos son los coanoflagelados, flagelados con la misma estructura que cierto tipo de células de las esponjas. Estudios moleculares los sitúan en el supergrupo de los opistocontos, que también incluye a los hongos y a pequeños protistas parasitarios emparentados con estos últimos. El nombre viene de la localización trasera del flagelo en las células móviles, como en muchos espermatozoides animales, mientras que otros eucariontes tienen flagelos delanteros (acrocontos).
Teoría simbióntica. Una segunda hipótesis contempla la posibilidad que diferentes Protistas se hubiesen asociado simbióticamente originando un organismo pluricelular. Este es el origen que se presupone para las células eucariotas a partir de células procariotas. No obstante, no hay pruebas que respalden el origen simbiótico de los metazoos.
Teoría de la celularización. Otra teoría, que provocó profundas divergencias entre los zoólogos, es la que contempla a los turbelarios como los metazoos más primitivos y por tanto cuestiona el carácter ancestral de cnidarios y esponjas. Según esta hipótesis, los turbelarios derivarían de protistas ciliados multinucleados, por medio de celularización de los núcleos, lo que concuerda con el concepto de protozoo como organismo acelular. No obstante, hay muchos aspectos en contra de esta teoría, ya que no tiene en cuenta los criterios fundamentados en la embriología y da mucha más importancia a la organización del adulto.
Los primeros fósiles que podrían representar animales aparecen hacia el final del Precámbrico, hace alrededor de 600 millones de años, y se les conoce como vendobiontes. Sin embargo, son muy difíciles de relacionar con los fósiles posteriores. Algunos de estos organismos podrían ser los precursores de los filos modernos, pero también podrían ser grupos separados, y es posible que no fueran realmente animales en sentido estricto. Aparte de ellos, muchos filos conocidos de animales hicieron una aparición más o menos simultánea durante el período Cámbrico, hace cerca de 570 millones de años. Todavía se discute si este evento, llamado explosión cámbrica, representa una rápida divergencia entre diferentes grupos o un cambio de condiciones que facilitó la fosilización.Entre los ancestros de grupos posteriores destacamos al Anomalocaris, del Cámbrico, como posible ancestro de diversos grupos posteriores de artrópodos, por su cuerpo segmentado, evolucionado de Opabinia y otros similares. Los cordados podrían tener relación con Pikaia
 |
| Amoeba |
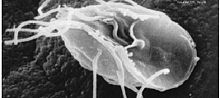 |
| Giarda,un flagelado parásito |
 |
| Un ciliado típico |
Los animales llevan a cabo las siguientes funciones esenciales: alimentación, respiración, circulación, excreción, respuesta, movimiento y reproducción:
Alimentación: La mayoría de los animales no pueden absorber comida; la ingieren. Los animales han evolucionado de diversas formas para alimentarse. Los herbívoros comen plantas, los carnívoros comen otros animales; y los omnívoros se alimentan tanto de plantas como de animales. Los detritívoros comen material vegetal y animal en descomposición. Los comedores por filtración son animales acuáticos que cuelan minúsculos organismos que flotan en el agua. Los animales también forman relaciones simbióticas, en las que dos especies viven en estrecha asociación mutua. Por ejemplo un parásito es un tipo de simbionte que vive dentro o sobre otro organismo, el huésped. El parásito se alimenta del huésped y lo daña.1
Respiración: no importa si viven en el agua o en la tierra, todos los animales respiran; esto significa que pueden tomar oxígeno y despedir dióxido de carbono. Gracias a sus cuerpos muy simples y de delgadas paredes, algunos animales utilizan la difusión de estas sustancias a través de la piel. Sin embargo, la mayoría de los animales han evolucionado complejos tejidos y sistemas orgánicos para la respiración.1
Circulación: Muchos animales acuáticos pequeños, como algunos gusanos, utilizan solo la difusión para transportar oxígeno y moléculas de nutrientes a todas sus células, y recoger de ellas los productos de desecho. La difusión basta porque estos animales apenas tienen un espesor de unas cuantas células. Sin embargo, los animales más grandes poseen algún tipo de sistema circulatorio para desplazar sustancias por el interior de sus cuerpos.1
Excreción: un producto de desecho primario de las células es el amoniaco, sustancia venenosa que contiene nitrógeno. La acumulación de amoniaco y otros productos de desecho podrían matar a un animal. La mayoría de los animales poseen un sistema excretor que bien elimina amoniaco o bien lo transforma en una sustancia menos tóxica que se elimina del cuerpo. Gracias a que eliminan los desechos metabólicos, los sistemas excretores ayudan a mantener la homeóstasis. Los sistemas excretores varían, desde células que bombean agua fuera del cuerpo hasta órganos complejos como riñones.
Respuesta: Los animales usan células especializadas, llamadas células nerviosas, para responder a los sucesos de su medio ambiente. En la mayoría de los animales, las células nerviosas eAunque no se dispone de todos los datos al completo, está claro que los dinosaurios no avianos, en general, eran voluminosos. Incluso dentro de sus propios estándares, los saurópodos alcanzaban el estatus de gigantescos: durante buena parte de su reinado, hasta los más pequeños de este subgrupo eran considerablemente mayores que cualquier otro animal de su hábitat, y los más grandes eran por lo menos un orden de magnitud mayores que cualquier otro vertebrado que caminara sobre la faz del planeta. Ésta es, posiblemente, una de las razones principales del interés que estas bestias han despertado en el público en general a través de los años. Pero la evolución también hizo aparecer dinosaurios pequeños, algunos del tamaño de una gallina, como Compsognathus, y más diminutos aún. El material fósil recuperado de estos "dinosaurios enanos" no suele ser tan completo, ya que sus huesos eran mucho más frágiles, lo que dificultó su preservación.
La mayoría de los saurópodos no podía desplazarse velozmente, debido a que tenían que soportar su colosal tamaño y peso. Existen límites prácticos al crecimiento: si estas criaturas se hubieran hecho más grandes hubieran quedado casi inmovilizadas y disfuncionales, lo que habría imposibilitado su supervivencia a largo plazo. Sin embargo, el hecho de que estas conformaciones gigantescas hayan sido viables durante millones de años, mucho más tiempo que el que han existido otras especies que consideramos exitosas, como el hombre, indica que estaban muy bien adaptadas a sus ambientes.
Esqueleto de Giraffatitan.
Por otro lado, existieron dinosaurios que fueron ágiles, delgados y pequeños. Particularmente, se considera a los tetanuros como los dinosaurios más rápidos; Tyrannosaurus, a pesar de ser uno de los tetanuros carnívoros más grandes, se desplazaba con cierta velocidad debido en parte a que su cráneo presentaba grandes órbitas en donde se situaban los ojos y nariz, con lo que su peso resultaba notablemente aligerado.
Una breve comparación con la fauna actual ilustra el dramático contraste de escalas: el mayor elefante registrado pesó 12 toneladas, mientras que la jirafa más alta alcanzó los 6 metros de altura. Incluso mamíferos prehistóricos gigantes como Indricotherium, o Mammuthus columbi quedan empequeñecidos por los saurópodos gigantes. Sólo unos pocos animales acuáticos modernos se les aproxima en tamaño, siendo el más notable la ballena azul, con un récord en el espécimen más grande registrado de 190 toneladas, y 33,5 metros de largo. Los dinosaurios no avianos más pequeños eran del tamaño de gallinas: Microraptor no superaba los 50 cm.
[editar]Extremos
El más alto y pesado de los dinosaurios de los cuales se recuperó el esqueleto completo fue Brachiosaurus, espécimen que fue descubierto en Tanzania entre 1907 y 1912. Se encuentra expuesto en el Museo Humboldt de Berlín, y tiene 12 metros de alto y probablemente haya pesado de 40 a 60 toneladas. El más largo de los hallados completos es Diplodocus, de unos 30 metros, recuperado en Wyoming, en los Estados Unidos y exhibido en el Museo de Historia Natural de Carnegie en 1907.
Estas cifras empalidecen cuando se las compara con nuevas especies halladas después de 1970, como Argentinosaurus, que podría haber pesado unas 100 toneladas; Seismosaurus, que habría alcanzado los 50 metros de largo; o Sauroposeidon, de unos 17 metros de altura.
Adicionalmente, el saurópodo más grande del que se tenga noticia parece haber sido Bruhathkayosaurus. Los vestigios fósiles exhumados parecen corresponder a un fémur, cadera, tibia, radio y una vértebra (todos en mal estado de conservación), aunque no se tiene seguridad de a qué clase de dinosaurio representan (la clasificación del espécimen ha sido singularmente errática), y si era en realidad un animal (estudios más detallados del material recuperado sugieren que se trata de restos vegetales). Asimismo el descubrimiento de otro posible gigante, Amphicoelias, ha sido objeto de fuerte escepticismo debido a la total ausencia de fósiles (la única prueba fue una supuesta vértebra de 2,5 metros que —según sus descubridores— se destruyó completamente durante el traslado). El hecho de que el anuncio del descubrimiento y sus irregulares condiciones ocurriera en medio de la llamada Guerra de los Huesos resta aún más confiabilidad a la versión de la existencia de la criatura.
[editar]Tamaño medio
Tamaño de un humano en comparación con el de Tyrannosaurus rex.
La distribución geográfica de los dinosaurios comenzó a finales del Triásico, produciéndose drásticas variaciones en forma, comportamiento y tamaño.
No obstante, el significado de "tamaño promedio" no es tan sencillo de precisar. Las observaciones actuales sugieren diferentes valores para cada uno de los períodos geológicos involucrados.2
De acuerdo con Bill Erickson:
"Los estimados de dinosaurios medianos oscilan entre 500 kg y 5 toneladas [...] Ochenta por ciento de la biomasa de la Formación de Morrison del oeste de los Estados Unidos consistía en Stegosaurus y saurópodos; este último promediaba las 20 toneladas [...]. El típico gran porte de los dinosaurios y el comparativamente pequeño tamaño de los mamíferos modernos ha sido cuantificado por Nicholas Hotton. Basándose en 63 géneros de dinosaurios, los datos de Hotton muestran una masa promedio que excede los 395.9 kg (el peso de un oso grizzly mediano), y una masa genérica mediana de cerca de dos toneladas (comparable a una jirafa). Esto contrasta marcadamente con mamíferos extintos (788 géneros) cuya masa promedio es de 631 gramos (la de un roedor pequeño). El dinosaurio no aviano más pequeño hallado hasta hoy era mayor que dos tercios de todos los mamíferos actuales; la mayoría de los dinosaurios eran mayores que todos los mamíferos vivientes con excepción de un 2% de los individuos".3
[editar]Comportamiento
Las interpretaciones sobre el comportamiento de los dinosaurios son inferencias generalmente basadas en la posición de los restos fósiles, en su hábitat, en simulaciones por computadora de la biomecánica de sus organismos, y en comparaciones con animales actuales de similares nichos ecológicos. Como resultado del carácter especulativo de estas fuentes, el entendimiento actual del comportamiento recae principalmente en la deducción científica y es a menudo controvertido. Sin embargo los científicos consideran que las actitudes propias de cocodrilos y aves, los seres vivos más cercanos a los dinosaurios, pueden ser extrapoladas en cierta medida con el fin de presentar un esquema de comportamiento posible para caracterizar al grupo.
[editar]Naturaleza gregaria
La primera prueba directa de comportamiento gregario la aportó en 1878 el descubrimiento en el yacimiento de Bernissart (Bélgica) de un yacimiento con 31 iguanodontes que habrían perecido juntos por ahogamiento al caer en una brecha profunda e inundada. El hallazgo de otros sitios que documentan muertes masivas y de pistas conteniendo abundantes huellas fosilizadas sugieren que el comportamiento de manada era común en varias especies de dinosaurios. Dichas huellas, a veces contadas de a miles, demuestran que géneros como Hadrosaurus, por ejemplo, podrían haberse movilizado en grandes grupos, tal como hoy lo hacen el bisonte americano o la gacela Springbok africana. Pisadas de saurópodos halladas en Oxford, Inglaterra, muestran que estos animales viajaban en grupos compuestos por diferentes especies.4 Un hallazgo realizado en Davenport Ranch, Texas, sugiere que algunos dinosaurios mantenían en el centro de la manada a los miembros juveniles para mayor defensa, posiblemente durante largas migraciones. [cita requerida]
Se ha especulado que el colosal tamaño de algunos saurópodos, sus formidables armas naturales, unidas a la protección adicional de la manada les podría haber conferido cierta invulnerabilidad, incluso frente a los terópodos más grandes. Sólo los individuos juveniles habrían sido presas potenciales de estos carnívoros. Por otro lado se cree que el desarrollo de estrategias cooperativas de caza podría haber facilitado a los atacantes el aislar a los adultos de las crías, lo que habría restaurado el equilibro poblacional.[cita requerida]
[editar]Patrones de anidación
El nido fosilizado de un Maiasaura con sus crías, descubierto en 1978.
El descubrimiento de Jack Horner en 1978 de un nido de Maiasaura ('atenta madre lagarto')5 en Montana, Estados Unidos, demostró que entre ornitópodos el cuidado paternal continuaba hasta mucho después del nacimiento.6 7
Existen asimismo pruebas de que otros dinosaurios del Cretácico, como el saurópodo patagónico Saltasaurus, tenían un comportamiento similar para el anidamiento, y que los individuos se agrupaban en inmensas colonias tal como hoy lo hacen los pingüinos.
El maniraptor Oviraptor que vivió en la actual Mongolia hace 65-70 millones de años, fue descubierto en 1993 en una posición similar a la de una gallina en acto de empollar, lo que puede implicar que estaba cubierto de alguna capa aislante de plumas (o tejido afín) que mantenía al huevo caliente.8 En su momento, se pensó que este singular terópodo de característicos cresta y pico habría tenido una alimentación basada en huevos.
Las marcas de pisadas también han confirmado un comportamiento paternal entre saurópodos y ornitópodos en la Isla de Skye, en el noroeste de Escocia.9
Nidos y huevos de los principales grupos de dinosaurios han sido descubiertos, y parece probable que los adultos hubieran tenido algún tipo de comunicación con sus crías, en forma similar a las aves actuales. [cita requerida]
[editar]Cortejo y apareamiento
Los adornos craneales de algunos dinosaurios, tales como crestas sagitales, cuernos, y demás protuberancias comunes en grupos como Marginocephalia, podrían haber sido demasiado frágiles como para haber desempeñado algún rol activo en la defensa, por lo que los investigadores consideran que tenían un uso meramente demostrativo, especialmente relacionado con actividades de tipo sexual. Se conoce muy poco sobre el apareamiento o el territorialismo de estos animales. La naturaleza de la comunicación social entre dinosaurios también permanece en las sombras, a pesar de que ambas son activas áreas de la investigación actual. Por ejemplo investigaciones recientes sugieren que las crestas huecas de los lambeosáuridos podrían haber funcionado como una cámara de resonancia con funciones de vocalización. [cita requerida]
Sin embargo, algunos dinosaurios herbívoros efectivamente presentaban a veces formidables y efectivas defensas corporales, tales como las grandes placas óseas de los estegosaurios, los cuernos de los ceratopsianos, o las corazas de espinas de los anquilosaurios. Algunas de estas características —desarrolladas durante el Mesozoico— quizá desempeñaron funciones secundarias, como la de regulación térmica y de lucimiento para el apareamiento: se considera que todos los ceratopsianos, entre ellos Triceratops, usaban sus cuernos para defenderse o pelear entre ellos durante el cortejo.
[editar]Depredación
Un Allosaurus devorando carroña de un saurópodo.
Desde el punto de vista del comportamiento, uno de los fósiles más valiosos fue desenterrado en el Desierto de Gobi en 1971. Incluía un Velociraptor atacando a un Protoceratops,10 lo que demostró la forma en que los dinosaurios se atacaban y defendían mutuamente. Mientras que actitudes canibalísticas entre terópodos no resultan sorprendentes,11 éstas también fueron confirmadas por marcas de dientes obtenidas en fósiles de Madagascar en 2003.12
Se presume que algunos terópodos cazaban en manada, como Velociraptor, mientras que carnívoros más grandes como Tyrannosaurus lo hacían en solitario (otros investigadores sostienen que este último grupo pudo haber sido principalmente carroñero) [cita requerida].
Una de las razones por las cuales se piensa que los dinosaurios fueron endotérmicos es que los fósiles en su conjunto parecen indicar que al menos los carnívoros fueron muy activos, desplegando mucha energía para perseguir y matar a sus presas. Las hipotéticas estrategias de caza de estos animales implican cierto desarrollo de la inteligencia, agilidad y fuerza, que hacen necesaria una fisiología más avanzada.[cita requerida]
[editar]Biomecánica
Existen pruebas de especies que trepaban árboles como Microraptor y también se tiene constancia de algunas que cavaron madrigueras como el hipsilofodóntido Oryctodromeus. Esto es bastante sorprendente si se tiene en cuenta que estas actividades eran comunes en los mamíferos del Cenozoico. Acerca de la fenomenología del movimiento, la biomecánica ha producido avances significativos. Por ejemplo, el estudio de las fuerzas ejercidas por los músculos y la gravedad en el esqueleto de estos animales mostraron cuan velozmente podían desplazarse.13 14
Existen sospechas de que algunos diplodócidos podrían haber creado estampidos sónicos mediante la ondulación de sus colas en forma de látigo.15 Sin embargo, esta afirmación está siendo investigada ya que los huesos de la cola serían demasiado frágiles para aguantar el gran impacto que supondría que actuasen como un látigo, y las vértebras se quebrarían, por lo que no resulta un comportamiento muy efectivo.
Finalmente, se ha investigado si los terópodos gigantes tenían que controlar su velocidad al arremeter contra sus presas para evitarse heridas fatales,16 y si los saurópodos podían flotar en agua
Fisiología
Esqueletos de dinosaurios expuestos en el Museo Real de Ontario, Canadá.
Los reptiles como los lagartos y serpientes son ectotérmicos (también llamados "de sangre fría"), es decir, obtienen la mayor parte de su calor corporal del Sol. Este tipo de fisiología requiere en general menos alimentos, agua y oxígeno para mantener al individuo activo. Estos animales son capaces de respuestas enérgicas aunque breves, caracterizadas por reacciones de súbita velocidad seguidas por períodos prolongados de reposo; es decir no pueden mantener una actividad vigorosa durante lapsos extensos. El consenso entre los estudiosos, inicialmente inclinado hacia la naturaleza ectotérmica, fue modificado tras los trabajos que Robert T. Bakker publicó en 1968. Hoy el punto de vista predominante favorece la versión endotérmica, o de "sangre caliente", basándose en pruebas recientes:
Las extremidades debajo del cuerpo de los dinosaurios sugieren un estilo de vida activo, particularmente en el caso de los terópodos.
Las crías de dinosaurios podían crecer rápidamente (esto se sabe gracias a estudios de desarrollo comparado en diversos géneros), como las de los mamíferos y las aves.
Hubo géneros cuya piel se hallaba cubierta por formaciones filamentosas muy similares a plumas, que podrían haber desempeñado un papel de aislamiento térmico. Útil para un animal endotermo pero que dificultaría el asoleamiento de un ectotermo.
La estructura física de los terópodos era semejante a la de las aves.
Hubo dinosaurios que habitaron en regiones semipolares, donde ningún animal ectotermo podría haber existido.
La conformación de los canales venosos presentes en el ánima ósea es muy similar a la de los mamíferos y aves actuales.
La complejidad del problema radica en el hecho que la endotermia puede desarrollarse por medio de más de un mecanismo. La mayoría de las discusiones sobre el tema tienden a comparar a los dinosaurios con los pájaros o los mamíferos, que gastan energía para elevar la temperatura del cuerpo. Aves y mamíferos pequeños añaden otro recurso: el aislamiento térmico, compuesto por tejidos como grasa, dermis gruesa, o plumas, que retrasan la pérdida de calor. Sin embargo, los mamíferos grandes, tales como los elefantes, hacen frente a otro problema debido a su cociente relativamente pequeño entre superficie y volumen (principio de Haldane). Este cociente relaciona el volumen de un animal con el área de su piel: al duplicarse las dimensiones de una criatura, su área superficial se cuadruplica mientras que el volumen se octuplica. En cierto punto, la cantidad de calor irradiado a través de la piel es superada por la cantidad de calor producida dentro del cuerpo por el metabolismo, forzando a estos grandes animales a utilizar métodos adicionales para evitar el recalentamiento. En el caso particular de los elefantes, han desarrollado diversas tácticas evolutivas como pérdida del pelo, extensos pabellones auriculares que aumentan el área superficial sin incrementar significativamente el volumen, y adaptaciones del comportamiento, como por ejemplo el rociar agua sobre sus propios cuerpos y practicar baños de fango refrescantes en forma periódica.
Los dinosaurios de grandes dimensiones deberían haber enfrentado una problemática similar. Muchos investigadores creen que son justamente estas grandes proporciones las que les habrían permitido conseguir propiedades endotérmicas al mantener templado el interior de sus cuerpos por simple acumulación pasiva de calor y sin contar con los mecanismos activos propios de criaturas más evolucionadas. Esta hipótesis, sin embargo, no puede explicar cómo los dinosaurios más pequeños —que formaban la mayoría del ecosistema— podían mantener los niveles de actividad que sus características físicas parecen insinuar, por lo que el debate continúa.
Modelo de Archaeopteryx lithographica basado en el fósil expuesto en el Museo de Historia Natural de la Universidad de Oxford.
Se ha probado sólidamente que existió una transición evolutiva desde ciertos dinosaurios hacia las aves. Las aves son claramente monofiléticas y sus primeros representantes se encuentran en el Jurásico (Protoavis, del Triásico, fue propuesto como ave pero se trata de un fósil de interpretación muy dudosa).
[editar]Plumas
Archaeopteryx, la primera ave, fue descubierta en 1861. El espécimen original fue encontrado en las calizas de Solnhofen en Alemania meridional, una destacable y poco común formación geológica conocida por sus extraordinariamente bien conservados fósiles. Archaeopteryx es un fósil transicional con características claramente atribuibles a modernos reptiles y a aves por igual. Hallado dos años después de la publicación de El origen de las especies de Charles Darwin, su descubrimiento y probable rol de "eslabón perdido" catapultó el debate entre biólogos evolutivos y creacionistas. Esta ave primitiva es tan similar a un dinosaurio que, cuando las plumas no estaban preservadas en improntas fósiles, sus ejemplares fueron confundidos con individuos del género Compsognathus.
Desde los años 1990 se ha encontrado un número creciente de dinosaurios no avianos con plumas, lo cual proveyó nuevas pruebas adicionales sobre la relación directa existente entre dinosaurios y pájaros. Los fósiles involucrados presentan rasgos de aves, incluyendo plumas que no son exactamente como las de las aves actuales, sino que reúnen algunas características intermedias entre el pelo y el plumón, útiles para cubrirse del frío. Este tipo de pluma, más conocida como "protopluma", es aparentemente la precursora del tipo que las aves modernas poseen. Entre los dinosaurios emplumados descubiertos hasta ahora están Beipiaosaurus, Caudipteryx, Dilong, Microraptor, Protarchaeopteryx, Shuvuuia, Sinornithosaurus, Sinosauropteryx y Jinfengopteryx. También se han encontrado huellas de dinosaurios terópodos, las cuales se asemejan llamativamente a improntas de aves. Entre los dromeosáuridos, los dinosaurios más próximos a las aves, se encuentra al menos un género alado y dotado de plumas asimétricas (aerodinámicas): el Microraptor. Otros dromeosáuridos posteriores, como el Velociraptor, podrían haber evolucionado a partir de formas con alas.
La mayoría de estos especímenes han sido exhumados en la provincia de Liaoning en el noreste de China, que fue parte de un continente aislado durante el Cretáceo. A pesar de que rastros de plumas han sido encontrados sólo en la formación Yixian y otros pocos sitios, es probable que dinosaurios similares en otras partes del mundo también hayan estado cubiertos por plumas. La falta de restos fósiles de plumaje a escala planetaria puede deberse al hecho de que las estructuras delicadas como piel, pelo y plumas no son preservadas por las usuales condiciones de fosilización, lo que dificulta su presencia en el registro fósil.
[editar]Esqueleto
Los dinosaurios y las aves comparten muchas características: se han identificado más de cien rasgos anatómicos aviares —especialmente esqueléticos— en común con los terópodos maniraptores, aceptados generalmente como sus parientes más cercanos. Algunos de estos rasgos en común se presentan en el cuello, pubis, muñeca, extremidades superiores, clavícula, omóplato, cerco y huesos pectorales. 19 Todo esto afirma la idea de que ciertos dinosaurios fueron efectivamente los antepasados de las aves, a cuya forma convergieron en un largo proceso de adaptaciones anatómicas y fisiológicas.
[editar]Similitudes reproductivas
Cráneo de Tyrannosaurus rex en el Museo de Historia Natural de Carnegie.
Un reciente descubrimiento efectuado en un esqueleto de Tyrannosaurus rex suministró pruebas adicionales del origen dinosauriano de las aves y, por primera vez, permitió a los paleontólogos establecer el sexo de un dinosaurio. Cuando depositan sus huevos, las aves hembras desarrollan un tipo especial de tejido óseo en sus extremidades, llamado hueso medular, rico en calcio, que forma una capa dentro del hueso duro exterior, y que sirve para formar la cáscara del huevo. La presencia de esta clase de tejido óseo en el tuétano de porciones de los restos de las patas traseras de Tyrannosaurus sugiere que este animal usó estrategias reproductivas similares.
[editar]Pulmones
Los grandes dinosaurios carnívoros tuvieron un sistema de sacos de aire similar al que se encuentra en las aves modernas, de acuerdo a una investigación dirigida por Patrick O'Connor, de la Universidad de Ohio. Los pulmones de los terópodos probablemente empujaban aire hacia sacos vacíos en sus esqueletos, como en el caso de las aves. "Lo que una vez fue formalmente considerado exclusivo de las aves estuvo presente de alguna forma en sus ancestros", declaró O'Connor. El estudio fue parcialmente financiado por la Fundación Nacional para la Ciencia estadounidense.20
[editar]El corazón y la postura durante el sueño
Análisis de cavidades pectorales de dinosaurios hechas con tomografía computarizada en 2000 revelaron aparentes remanentes de complejos corazones de cuatro cavidades, parecidos a los que hoy tienen los mamíferos y las aves. Un fósil de Troodon recientemente descubierto demuestra que estos dinosaurios durmieron como ciertas aves actuales, con sus cabezas escondidas bajo los brazos, postura que ayuda a conservar el calor craneal.21 Este comportamiento pudo haber ayudado a mantener la cabeza caliente.
[editar]Gastrolitos
Otra prueba de que los dinosaurios y las aves están estrechamente relacionados es la ingestión de piedras con fines digestivos, las que son tragadas por estos animales y ayudan a triturar las fibras resistentes de la comida cuando entran al estómago. Encontradas en asociación con fósiles, estas rocas son denominadas gastrolitos. Debido a que piedras identificables como pertenecientes a cierta formación geológica pueden haber sido engullidas en un lugar y trasladadas dentro del animal en sus desplazamientos, los paleontólogos a veces las usan para establecer posibles rutas de migración.
[editar]Tejido blando en fósiles de dinosaurios
Uno de los mejores ejemplos de impresiones fósiles de tejido blando de dinosaurio se descubrió en Petraroia, Italia. El hallazgo fue informado en 1998, y la huella se describió como "dejada por un espécimen pequeño", un muy joven coelurosáurido, Scipionyx samniticus. El fósil incluye porciones de intestinos, colon, hígado, músculos y tráquea de este dinosaurio inmaduro.
En el número de marzo de 2005 de la revista Science, la Dra. Mary Higby Schweitzer y su equipo anunciaron el descubrimiento de una materia flexible que parece ser tejido suave de la pierna de un Tyrannosaurus rex de 68 millones de años de antigüedad en la formación del Riachuelo del Infierno, en Montana, Estados Unidos. Después de la recuperación, el tejido fue rehidratado por el equipo científico.
Cuando el hueso fosilizado fue tratado durante varias semanas para separar el contenido mineral de la cavidad del tuétano (un proceso llamado desmineralización), Schweitzer encontró pruebas de estructuras intactas como vasos sanguíneos, matriz ósea y tejido conectivo (fibras óseas). Analizado bajo el microscopio, el tejido reveló contener microestructuras intactas incluso hasta el nivel celular. La naturaleza y composición exactas del material no están todavía claras, aunque muchas notas de prensa exageradamente relacionaron el hallazgo con el tema de la novela Parque Jurásico. La interpretación del descubrimiento, así como su importancia relativa, todavía están siendo discutidas.


La mayoría de los saurópodos no podía desplazarse velozmente, debido a que tenían que soportar su colosal tamaño y peso. Existen límites prácticos al crecimiento: si estas criaturas se hubieran hecho más grandes hubieran quedado casi inmovilizadas y disfuncionales, lo que habría imposibilitado su supervivencia a largo plazo. Sin embargo, el hecho de que estas conformaciones gigantescas hayan sido viables durante millones de años, mucho más tiempo que el que han existido otras especies que consideramos exitosas, como el hombre, indica que estaban muy bien adaptadas a sus ambientes.
Esqueleto de Giraffatitan.
Por otro lado, existieron dinosaurios que fueron ágiles, delgados y pequeños. Particularmente, se considera a los tetanuros como los dinosaurios más rápidos; Tyrannosaurus, a pesar de ser uno de los tetanuros carnívoros más grandes, se desplazaba con cierta velocidad debido en parte a que su cráneo presentaba grandes órbitas en donde se situaban los ojos y nariz, con lo que su peso resultaba notablemente aligerado.
Una breve comparación con la fauna actual ilustra el dramático contraste de escalas: el mayor elefante registrado pesó 12 toneladas, mientras que la jirafa más alta alcanzó los 6 metros de altura. Incluso mamíferos prehistóricos gigantes como Indricotherium, o Mammuthus columbi quedan empequeñecidos por los saurópodos gigantes. Sólo unos pocos animales acuáticos modernos se les aproxima en tamaño, siendo el más notable la ballena azul, con un récord en el espécimen más grande registrado de 190 toneladas, y 33,5 metros de largo. Los dinosaurios no avianos más pequeños eran del tamaño de gallinas: Microraptor no superaba los 50 cm.
[editar]Extremos
El más alto y pesado de los dinosaurios de los cuales se recuperó el esqueleto completo fue Brachiosaurus, espécimen que fue descubierto en Tanzania entre 1907 y 1912. Se encuentra expuesto en el Museo Humboldt de Berlín, y tiene 12 metros de alto y probablemente haya pesado de 40 a 60 toneladas. El más largo de los hallados completos es Diplodocus, de unos 30 metros, recuperado en Wyoming, en los Estados Unidos y exhibido en el Museo de Historia Natural de Carnegie en 1907.
Estas cifras empalidecen cuando se las compara con nuevas especies halladas después de 1970, como Argentinosaurus, que podría haber pesado unas 100 toneladas; Seismosaurus, que habría alcanzado los 50 metros de largo; o Sauroposeidon, de unos 17 metros de altura.
Adicionalmente, el saurópodo más grande del que se tenga noticia parece haber sido Bruhathkayosaurus. Los vestigios fósiles exhumados parecen corresponder a un fémur, cadera, tibia, radio y una vértebra (todos en mal estado de conservación), aunque no se tiene seguridad de a qué clase de dinosaurio representan (la clasificación del espécimen ha sido singularmente errática), y si era en realidad un animal (estudios más detallados del material recuperado sugieren que se trata de restos vegetales). Asimismo el descubrimiento de otro posible gigante, Amphicoelias, ha sido objeto de fuerte escepticismo debido a la total ausencia de fósiles (la única prueba fue una supuesta vértebra de 2,5 metros que —según sus descubridores— se destruyó completamente durante el traslado). El hecho de que el anuncio del descubrimiento y sus irregulares condiciones ocurriera en medio de la llamada Guerra de los Huesos resta aún más confiabilidad a la versión de la existencia de la criatura.
[editar]Tamaño medio
Tamaño de un humano en comparación con el de Tyrannosaurus rex.
La distribución geográfica de los dinosaurios comenzó a finales del Triásico, produciéndose drásticas variaciones en forma, comportamiento y tamaño.
No obstante, el significado de "tamaño promedio" no es tan sencillo de precisar. Las observaciones actuales sugieren diferentes valores para cada uno de los períodos geológicos involucrados.2
De acuerdo con Bill Erickson:
"Los estimados de dinosaurios medianos oscilan entre 500 kg y 5 toneladas [...] Ochenta por ciento de la biomasa de la Formación de Morrison del oeste de los Estados Unidos consistía en Stegosaurus y saurópodos; este último promediaba las 20 toneladas [...]. El típico gran porte de los dinosaurios y el comparativamente pequeño tamaño de los mamíferos modernos ha sido cuantificado por Nicholas Hotton. Basándose en 63 géneros de dinosaurios, los datos de Hotton muestran una masa promedio que excede los 395.9 kg (el peso de un oso grizzly mediano), y una masa genérica mediana de cerca de dos toneladas (comparable a una jirafa). Esto contrasta marcadamente con mamíferos extintos (788 géneros) cuya masa promedio es de 631 gramos (la de un roedor pequeño). El dinosaurio no aviano más pequeño hallado hasta hoy era mayor que dos tercios de todos los mamíferos actuales; la mayoría de los dinosaurios eran mayores que todos los mamíferos vivientes con excepción de un 2% de los individuos".3
[editar]Comportamiento
Las interpretaciones sobre el comportamiento de los dinosaurios son inferencias generalmente basadas en la posición de los restos fósiles, en su hábitat, en simulaciones por computadora de la biomecánica de sus organismos, y en comparaciones con animales actuales de similares nichos ecológicos. Como resultado del carácter especulativo de estas fuentes, el entendimiento actual del comportamiento recae principalmente en la deducción científica y es a menudo controvertido. Sin embargo los científicos consideran que las actitudes propias de cocodrilos y aves, los seres vivos más cercanos a los dinosaurios, pueden ser extrapoladas en cierta medida con el fin de presentar un esquema de comportamiento posible para caracterizar al grupo.
[editar]Naturaleza gregaria
La primera prueba directa de comportamiento gregario la aportó en 1878 el descubrimiento en el yacimiento de Bernissart (Bélgica) de un yacimiento con 31 iguanodontes que habrían perecido juntos por ahogamiento al caer en una brecha profunda e inundada. El hallazgo de otros sitios que documentan muertes masivas y de pistas conteniendo abundantes huellas fosilizadas sugieren que el comportamiento de manada era común en varias especies de dinosaurios. Dichas huellas, a veces contadas de a miles, demuestran que géneros como Hadrosaurus, por ejemplo, podrían haberse movilizado en grandes grupos, tal como hoy lo hacen el bisonte americano o la gacela Springbok africana. Pisadas de saurópodos halladas en Oxford, Inglaterra, muestran que estos animales viajaban en grupos compuestos por diferentes especies.4 Un hallazgo realizado en Davenport Ranch, Texas, sugiere que algunos dinosaurios mantenían en el centro de la manada a los miembros juveniles para mayor defensa, posiblemente durante largas migraciones. [cita requerida]
Se ha especulado que el colosal tamaño de algunos saurópodos, sus formidables armas naturales, unidas a la protección adicional de la manada les podría haber conferido cierta invulnerabilidad, incluso frente a los terópodos más grandes. Sólo los individuos juveniles habrían sido presas potenciales de estos carnívoros. Por otro lado se cree que el desarrollo de estrategias cooperativas de caza podría haber facilitado a los atacantes el aislar a los adultos de las crías, lo que habría restaurado el equilibro poblacional.[cita requerida]
[editar]Patrones de anidación
El nido fosilizado de un Maiasaura con sus crías, descubierto en 1978.
El descubrimiento de Jack Horner en 1978 de un nido de Maiasaura ('atenta madre lagarto')5 en Montana, Estados Unidos, demostró que entre ornitópodos el cuidado paternal continuaba hasta mucho después del nacimiento.6 7
Existen asimismo pruebas de que otros dinosaurios del Cretácico, como el saurópodo patagónico Saltasaurus, tenían un comportamiento similar para el anidamiento, y que los individuos se agrupaban en inmensas colonias tal como hoy lo hacen los pingüinos.
El maniraptor Oviraptor que vivió en la actual Mongolia hace 65-70 millones de años, fue descubierto en 1993 en una posición similar a la de una gallina en acto de empollar, lo que puede implicar que estaba cubierto de alguna capa aislante de plumas (o tejido afín) que mantenía al huevo caliente.8 En su momento, se pensó que este singular terópodo de característicos cresta y pico habría tenido una alimentación basada en huevos.
Las marcas de pisadas también han confirmado un comportamiento paternal entre saurópodos y ornitópodos en la Isla de Skye, en el noroeste de Escocia.9
Nidos y huevos de los principales grupos de dinosaurios han sido descubiertos, y parece probable que los adultos hubieran tenido algún tipo de comunicación con sus crías, en forma similar a las aves actuales. [cita requerida]
[editar]Cortejo y apareamiento
Los adornos craneales de algunos dinosaurios, tales como crestas sagitales, cuernos, y demás protuberancias comunes en grupos como Marginocephalia, podrían haber sido demasiado frágiles como para haber desempeñado algún rol activo en la defensa, por lo que los investigadores consideran que tenían un uso meramente demostrativo, especialmente relacionado con actividades de tipo sexual. Se conoce muy poco sobre el apareamiento o el territorialismo de estos animales. La naturaleza de la comunicación social entre dinosaurios también permanece en las sombras, a pesar de que ambas son activas áreas de la investigación actual. Por ejemplo investigaciones recientes sugieren que las crestas huecas de los lambeosáuridos podrían haber funcionado como una cámara de resonancia con funciones de vocalización. [cita requerida]
Sin embargo, algunos dinosaurios herbívoros efectivamente presentaban a veces formidables y efectivas defensas corporales, tales como las grandes placas óseas de los estegosaurios, los cuernos de los ceratopsianos, o las corazas de espinas de los anquilosaurios. Algunas de estas características —desarrolladas durante el Mesozoico— quizá desempeñaron funciones secundarias, como la de regulación térmica y de lucimiento para el apareamiento: se considera que todos los ceratopsianos, entre ellos Triceratops, usaban sus cuernos para defenderse o pelear entre ellos durante el cortejo.
[editar]Depredación
Un Allosaurus devorando carroña de un saurópodo.
Desde el punto de vista del comportamiento, uno de los fósiles más valiosos fue desenterrado en el Desierto de Gobi en 1971. Incluía un Velociraptor atacando a un Protoceratops,10 lo que demostró la forma en que los dinosaurios se atacaban y defendían mutuamente. Mientras que actitudes canibalísticas entre terópodos no resultan sorprendentes,11 éstas también fueron confirmadas por marcas de dientes obtenidas en fósiles de Madagascar en 2003.12
Se presume que algunos terópodos cazaban en manada, como Velociraptor, mientras que carnívoros más grandes como Tyrannosaurus lo hacían en solitario (otros investigadores sostienen que este último grupo pudo haber sido principalmente carroñero) [cita requerida].
Una de las razones por las cuales se piensa que los dinosaurios fueron endotérmicos es que los fósiles en su conjunto parecen indicar que al menos los carnívoros fueron muy activos, desplegando mucha energía para perseguir y matar a sus presas. Las hipotéticas estrategias de caza de estos animales implican cierto desarrollo de la inteligencia, agilidad y fuerza, que hacen necesaria una fisiología más avanzada.[cita requerida]
[editar]Biomecánica
Existen pruebas de especies que trepaban árboles como Microraptor y también se tiene constancia de algunas que cavaron madrigueras como el hipsilofodóntido Oryctodromeus. Esto es bastante sorprendente si se tiene en cuenta que estas actividades eran comunes en los mamíferos del Cenozoico. Acerca de la fenomenología del movimiento, la biomecánica ha producido avances significativos. Por ejemplo, el estudio de las fuerzas ejercidas por los músculos y la gravedad en el esqueleto de estos animales mostraron cuan velozmente podían desplazarse.13 14
Existen sospechas de que algunos diplodócidos podrían haber creado estampidos sónicos mediante la ondulación de sus colas en forma de látigo.15 Sin embargo, esta afirmación está siendo investigada ya que los huesos de la cola serían demasiado frágiles para aguantar el gran impacto que supondría que actuasen como un látigo, y las vértebras se quebrarían, por lo que no resulta un comportamiento muy efectivo.
Finalmente, se ha investigado si los terópodos gigantes tenían que controlar su velocidad al arremeter contra sus presas para evitarse heridas fatales,16 y si los saurópodos podían flotar en agua
Fisiología
Esqueletos de dinosaurios expuestos en el Museo Real de Ontario, Canadá.
Los reptiles como los lagartos y serpientes son ectotérmicos (también llamados "de sangre fría"), es decir, obtienen la mayor parte de su calor corporal del Sol. Este tipo de fisiología requiere en general menos alimentos, agua y oxígeno para mantener al individuo activo. Estos animales son capaces de respuestas enérgicas aunque breves, caracterizadas por reacciones de súbita velocidad seguidas por períodos prolongados de reposo; es decir no pueden mantener una actividad vigorosa durante lapsos extensos. El consenso entre los estudiosos, inicialmente inclinado hacia la naturaleza ectotérmica, fue modificado tras los trabajos que Robert T. Bakker publicó en 1968. Hoy el punto de vista predominante favorece la versión endotérmica, o de "sangre caliente", basándose en pruebas recientes:
Las extremidades debajo del cuerpo de los dinosaurios sugieren un estilo de vida activo, particularmente en el caso de los terópodos.
Las crías de dinosaurios podían crecer rápidamente (esto se sabe gracias a estudios de desarrollo comparado en diversos géneros), como las de los mamíferos y las aves.
Hubo géneros cuya piel se hallaba cubierta por formaciones filamentosas muy similares a plumas, que podrían haber desempeñado un papel de aislamiento térmico. Útil para un animal endotermo pero que dificultaría el asoleamiento de un ectotermo.
La estructura física de los terópodos era semejante a la de las aves.
Hubo dinosaurios que habitaron en regiones semipolares, donde ningún animal ectotermo podría haber existido.
La conformación de los canales venosos presentes en el ánima ósea es muy similar a la de los mamíferos y aves actuales.
La complejidad del problema radica en el hecho que la endotermia puede desarrollarse por medio de más de un mecanismo. La mayoría de las discusiones sobre el tema tienden a comparar a los dinosaurios con los pájaros o los mamíferos, que gastan energía para elevar la temperatura del cuerpo. Aves y mamíferos pequeños añaden otro recurso: el aislamiento térmico, compuesto por tejidos como grasa, dermis gruesa, o plumas, que retrasan la pérdida de calor. Sin embargo, los mamíferos grandes, tales como los elefantes, hacen frente a otro problema debido a su cociente relativamente pequeño entre superficie y volumen (principio de Haldane). Este cociente relaciona el volumen de un animal con el área de su piel: al duplicarse las dimensiones de una criatura, su área superficial se cuadruplica mientras que el volumen se octuplica. En cierto punto, la cantidad de calor irradiado a través de la piel es superada por la cantidad de calor producida dentro del cuerpo por el metabolismo, forzando a estos grandes animales a utilizar métodos adicionales para evitar el recalentamiento. En el caso particular de los elefantes, han desarrollado diversas tácticas evolutivas como pérdida del pelo, extensos pabellones auriculares que aumentan el área superficial sin incrementar significativamente el volumen, y adaptaciones del comportamiento, como por ejemplo el rociar agua sobre sus propios cuerpos y practicar baños de fango refrescantes en forma periódica.
Los dinosaurios de grandes dimensiones deberían haber enfrentado una problemática similar. Muchos investigadores creen que son justamente estas grandes proporciones las que les habrían permitido conseguir propiedades endotérmicas al mantener templado el interior de sus cuerpos por simple acumulación pasiva de calor y sin contar con los mecanismos activos propios de criaturas más evolucionadas. Esta hipótesis, sin embargo, no puede explicar cómo los dinosaurios más pequeños —que formaban la mayoría del ecosistema— podían mantener los niveles de actividad que sus características físicas parecen insinuar, por lo que el debate continúa.
Modelo de Archaeopteryx lithographica basado en el fósil expuesto en el Museo de Historia Natural de la Universidad de Oxford.
Se ha probado sólidamente que existió una transición evolutiva desde ciertos dinosaurios hacia las aves. Las aves son claramente monofiléticas y sus primeros representantes se encuentran en el Jurásico (Protoavis, del Triásico, fue propuesto como ave pero se trata de un fósil de interpretación muy dudosa).
[editar]Plumas
Archaeopteryx, la primera ave, fue descubierta en 1861. El espécimen original fue encontrado en las calizas de Solnhofen en Alemania meridional, una destacable y poco común formación geológica conocida por sus extraordinariamente bien conservados fósiles. Archaeopteryx es un fósil transicional con características claramente atribuibles a modernos reptiles y a aves por igual. Hallado dos años después de la publicación de El origen de las especies de Charles Darwin, su descubrimiento y probable rol de "eslabón perdido" catapultó el debate entre biólogos evolutivos y creacionistas. Esta ave primitiva es tan similar a un dinosaurio que, cuando las plumas no estaban preservadas en improntas fósiles, sus ejemplares fueron confundidos con individuos del género Compsognathus.
Desde los años 1990 se ha encontrado un número creciente de dinosaurios no avianos con plumas, lo cual proveyó nuevas pruebas adicionales sobre la relación directa existente entre dinosaurios y pájaros. Los fósiles involucrados presentan rasgos de aves, incluyendo plumas que no son exactamente como las de las aves actuales, sino que reúnen algunas características intermedias entre el pelo y el plumón, útiles para cubrirse del frío. Este tipo de pluma, más conocida como "protopluma", es aparentemente la precursora del tipo que las aves modernas poseen. Entre los dinosaurios emplumados descubiertos hasta ahora están Beipiaosaurus, Caudipteryx, Dilong, Microraptor, Protarchaeopteryx, Shuvuuia, Sinornithosaurus, Sinosauropteryx y Jinfengopteryx. También se han encontrado huellas de dinosaurios terópodos, las cuales se asemejan llamativamente a improntas de aves. Entre los dromeosáuridos, los dinosaurios más próximos a las aves, se encuentra al menos un género alado y dotado de plumas asimétricas (aerodinámicas): el Microraptor. Otros dromeosáuridos posteriores, como el Velociraptor, podrían haber evolucionado a partir de formas con alas.
La mayoría de estos especímenes han sido exhumados en la provincia de Liaoning en el noreste de China, que fue parte de un continente aislado durante el Cretáceo. A pesar de que rastros de plumas han sido encontrados sólo en la formación Yixian y otros pocos sitios, es probable que dinosaurios similares en otras partes del mundo también hayan estado cubiertos por plumas. La falta de restos fósiles de plumaje a escala planetaria puede deberse al hecho de que las estructuras delicadas como piel, pelo y plumas no son preservadas por las usuales condiciones de fosilización, lo que dificulta su presencia en el registro fósil.
[editar]Esqueleto
Los dinosaurios y las aves comparten muchas características: se han identificado más de cien rasgos anatómicos aviares —especialmente esqueléticos— en común con los terópodos maniraptores, aceptados generalmente como sus parientes más cercanos. Algunos de estos rasgos en común se presentan en el cuello, pubis, muñeca, extremidades superiores, clavícula, omóplato, cerco y huesos pectorales. 19 Todo esto afirma la idea de que ciertos dinosaurios fueron efectivamente los antepasados de las aves, a cuya forma convergieron en un largo proceso de adaptaciones anatómicas y fisiológicas.
[editar]Similitudes reproductivas
Cráneo de Tyrannosaurus rex en el Museo de Historia Natural de Carnegie.
Un reciente descubrimiento efectuado en un esqueleto de Tyrannosaurus rex suministró pruebas adicionales del origen dinosauriano de las aves y, por primera vez, permitió a los paleontólogos establecer el sexo de un dinosaurio. Cuando depositan sus huevos, las aves hembras desarrollan un tipo especial de tejido óseo en sus extremidades, llamado hueso medular, rico en calcio, que forma una capa dentro del hueso duro exterior, y que sirve para formar la cáscara del huevo. La presencia de esta clase de tejido óseo en el tuétano de porciones de los restos de las patas traseras de Tyrannosaurus sugiere que este animal usó estrategias reproductivas similares.
[editar]Pulmones
Los grandes dinosaurios carnívoros tuvieron un sistema de sacos de aire similar al que se encuentra en las aves modernas, de acuerdo a una investigación dirigida por Patrick O'Connor, de la Universidad de Ohio. Los pulmones de los terópodos probablemente empujaban aire hacia sacos vacíos en sus esqueletos, como en el caso de las aves. "Lo que una vez fue formalmente considerado exclusivo de las aves estuvo presente de alguna forma en sus ancestros", declaró O'Connor. El estudio fue parcialmente financiado por la Fundación Nacional para la Ciencia estadounidense.20
[editar]El corazón y la postura durante el sueño
Análisis de cavidades pectorales de dinosaurios hechas con tomografía computarizada en 2000 revelaron aparentes remanentes de complejos corazones de cuatro cavidades, parecidos a los que hoy tienen los mamíferos y las aves. Un fósil de Troodon recientemente descubierto demuestra que estos dinosaurios durmieron como ciertas aves actuales, con sus cabezas escondidas bajo los brazos, postura que ayuda a conservar el calor craneal.21 Este comportamiento pudo haber ayudado a mantener la cabeza caliente.
[editar]Gastrolitos
Otra prueba de que los dinosaurios y las aves están estrechamente relacionados es la ingestión de piedras con fines digestivos, las que son tragadas por estos animales y ayudan a triturar las fibras resistentes de la comida cuando entran al estómago. Encontradas en asociación con fósiles, estas rocas son denominadas gastrolitos. Debido a que piedras identificables como pertenecientes a cierta formación geológica pueden haber sido engullidas en un lugar y trasladadas dentro del animal en sus desplazamientos, los paleontólogos a veces las usan para establecer posibles rutas de migración.
[editar]Tejido blando en fósiles de dinosaurios
Uno de los mejores ejemplos de impresiones fósiles de tejido blando de dinosaurio se descubrió en Petraroia, Italia. El hallazgo fue informado en 1998, y la huella se describió como "dejada por un espécimen pequeño", un muy joven coelurosáurido, Scipionyx samniticus. El fósil incluye porciones de intestinos, colon, hígado, músculos y tráquea de este dinosaurio inmaduro.
En el número de marzo de 2005 de la revista Science, la Dra. Mary Higby Schweitzer y su equipo anunciaron el descubrimiento de una materia flexible que parece ser tejido suave de la pierna de un Tyrannosaurus rex de 68 millones de años de antigüedad en la formación del Riachuelo del Infierno, en Montana, Estados Unidos. Después de la recuperación, el tejido fue rehidratado por el equipo científico.
Cuando el hueso fosilizado fue tratado durante varias semanas para separar el contenido mineral de la cavidad del tuétano (un proceso llamado desmineralización), Schweitzer encontró pruebas de estructuras intactas como vasos sanguíneos, matriz ósea y tejido conectivo (fibras óseas). Analizado bajo el microscopio, el tejido reveló contener microestructuras intactas incluso hasta el nivel celular. La naturaleza y composición exactas del material no están todavía claras, aunque muchas notas de prensa exageradamente relacionaron el hallazgo con el tema de la novela Parque Jurásico. La interpretación del descubrimiento, así como su importancia relativa, todavía están siendo discutidas.
No hay comentarios:
Publicar un comentario